.svg) National Institute of General Medical Sciences |
 |
|
National Biomedical Resource for |
| Recent Highlights | |
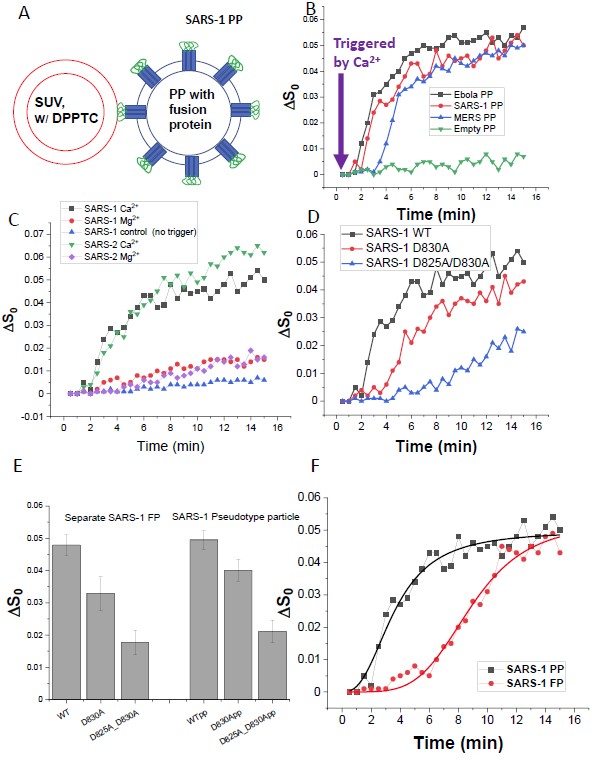
Entry of coronaviruses into host cells is mediated by the viral spike (S) protein. Previously, we identified the bona fide FPs for SARS-CoV ("SARS-1") and SARS-CoV-2 ("SARS-2") using ESR spectroscopy. We also found that their FPs induce membrane ordering in a Ca2+-dependent fashion. Here we study which negatively charged residues in SARS-1 FP are involved in this binding, to build a topological model and clarify the role of Ca2+. Our systematic mutation study on the SARS-1 FP shows that all six negatively charged residues contribute to the FP's membrane ordering activity, with D812 the dominant residue. The corresponding SARS-2 residue D830 plays an equivalent role. We provide a topological model of how the FP binds Ca2+ ions: its two segments FP1 and FP2 each bind one Ca2+. The binding of Ca2+, the folding of FP (both studied by ITC experiments), and the ordering activity correlate very well across the mutants, suggesting that the Ca2+ helps the folding of FP in membranes to enhance the ordering activity. Using a novel pseudotyped virus particle (PP)-liposome methodology, we monitored the membrane ordering induced by the FPs in the whole S protein in its trimer form in real time. We found that the SARS-1 and SARS-2 PPs also induce membrane ordering to the extent that separate FPs do, and mutations of the negatively charged residues also significantly suppress the membrane ordering activity. However, the slower kinetics of the FP ordering activity vs. that of the PP suggests the need for initial trimerization of the FPs. Publication: Biophys. J. 121, 207-227 (2022); PMC8683214. |
|
|
|
| Alex L. Lai, Jack H. Freed. (ACERT) | |
|
|
About ACERT Contact Us |
Research |
Outreach |
ACERT is supported by grant 1R24GM146107 from the National Institute of General Medical Sciences (NIGMS), part of the National Institutes of Health. |
|||||
| ||||||||