.svg) National Institute of General Medical Sciences |
 |
|
National Biomedical Resource for |
| ACERT's Service and Collaborative Projects | |
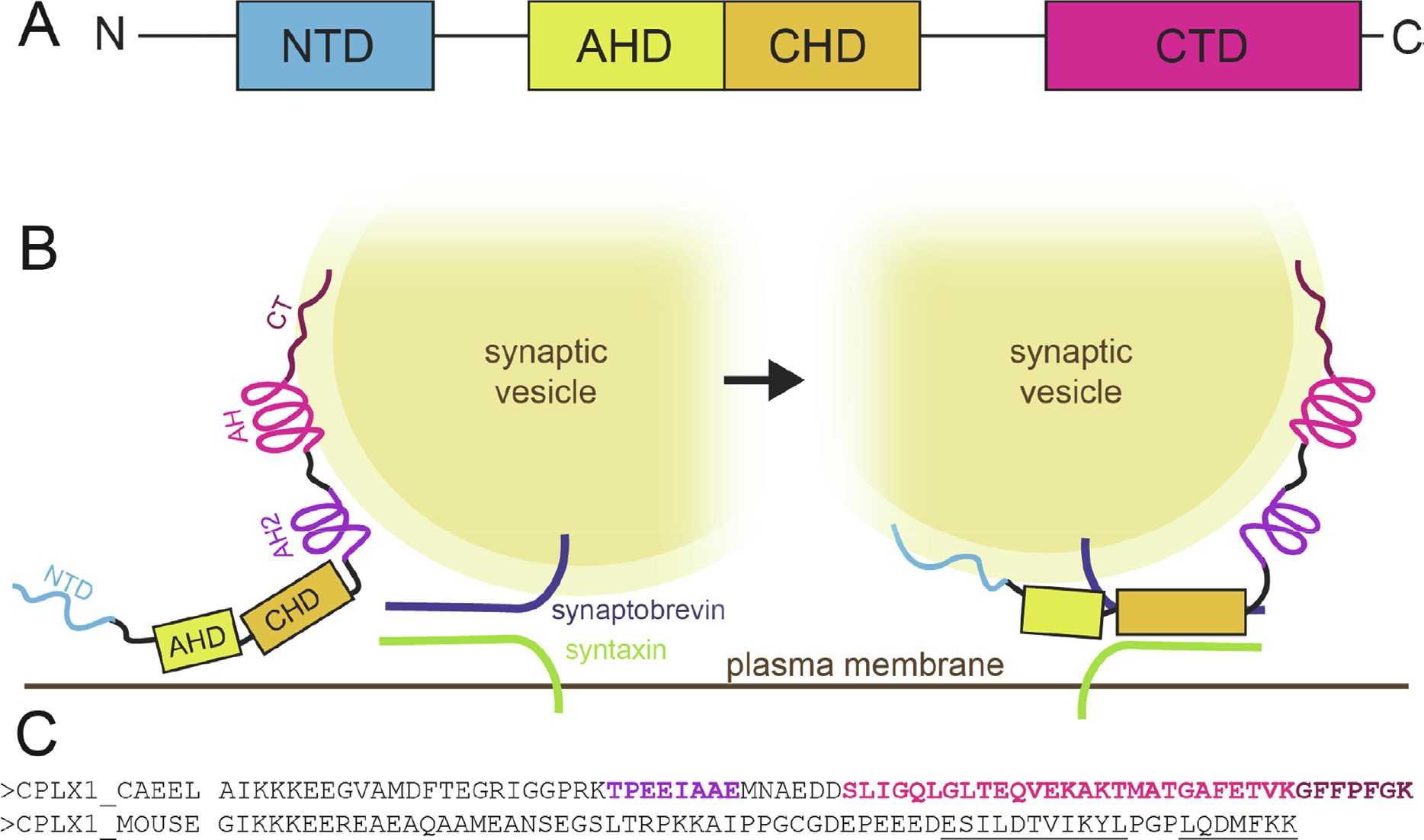
Complexins play a critical role in regulating SNARE-mediated exocytosis of synaptic vesicles. Evolutionary divergences in complexin function have complicated our understanding of the role these proteins play in inhibiting the spontaneous fusion of vesicles. Previous structural and functional characterizations of worm and mouse complexins have indicated the membrane curvature-sensing C-terminal domain of these proteins is responsible for differences in inhibitory function. We have characterized the structure and dynamics of the mCpx1 CTD in the absence and presence of membranes and membrane mimetics using NMR, ESR, and optical spectroscopies. In the absence of lipids, the mCpx1 CTD features a short helix near its N-terminus and is otherwise disordered. In the presence of micelles and small unilamellar vesicles, the mCpx1 CTD forms a discontinuous helical structure in its C-terminal 20 amino acids, with no preference for specific lipid compositions. In contrast, the mCpx1 CTD shows distinct compositional preferences in its interactions with large unilamellar vesicles. These studies identify structural divergences in the mCpx1 CTD relative to the wCpx1 CTD in regions that are known to be critical to the wCpx1 CTD's role in inhibiting spontaneous fusion of synaptic vesicles, suggesting a potential structural basis for evolutionary divergences in complexin function. Publication: J. Mol. Biol. 435 (1), 167710 (2023) PMC9794636 doi: 10.1016/j.jmb.2022.167710 |
|
|
|
|
Emily M. Grasso, Mayu S. Terakawa (Department of Biochemistry, Weill Cornell Medicine, New York, NY) Alex L. Lai (Department of Chemistry and Chemical Biology, Cornell University, Ithaca, NY) Ying Xue Xie, Trudy F. Ramlall (Department of Biochemistry, Weill Cornell Medicine, New York, NY) Jack H. Freed (Department of Chemistry and Chemical Biology, Cornell University, Ithaca, NY) and David Eliezer (Department of Biochemistry, Weill Cornell Medicine, New York, NY) |
|
|
|
About ACERT Contact Us |
Research |
Outreach |
ACERT is supported by grant 1R24GM146107 from the National Institute of General Medical Sciences (NIGMS), part of the National Institutes of Health. |
|||||
| ||||||||