.svg) National Institute of General Medical Sciences |
 |
|
National Biomedical Resource for |
| ACERT's Service and Collaborative Projects | |
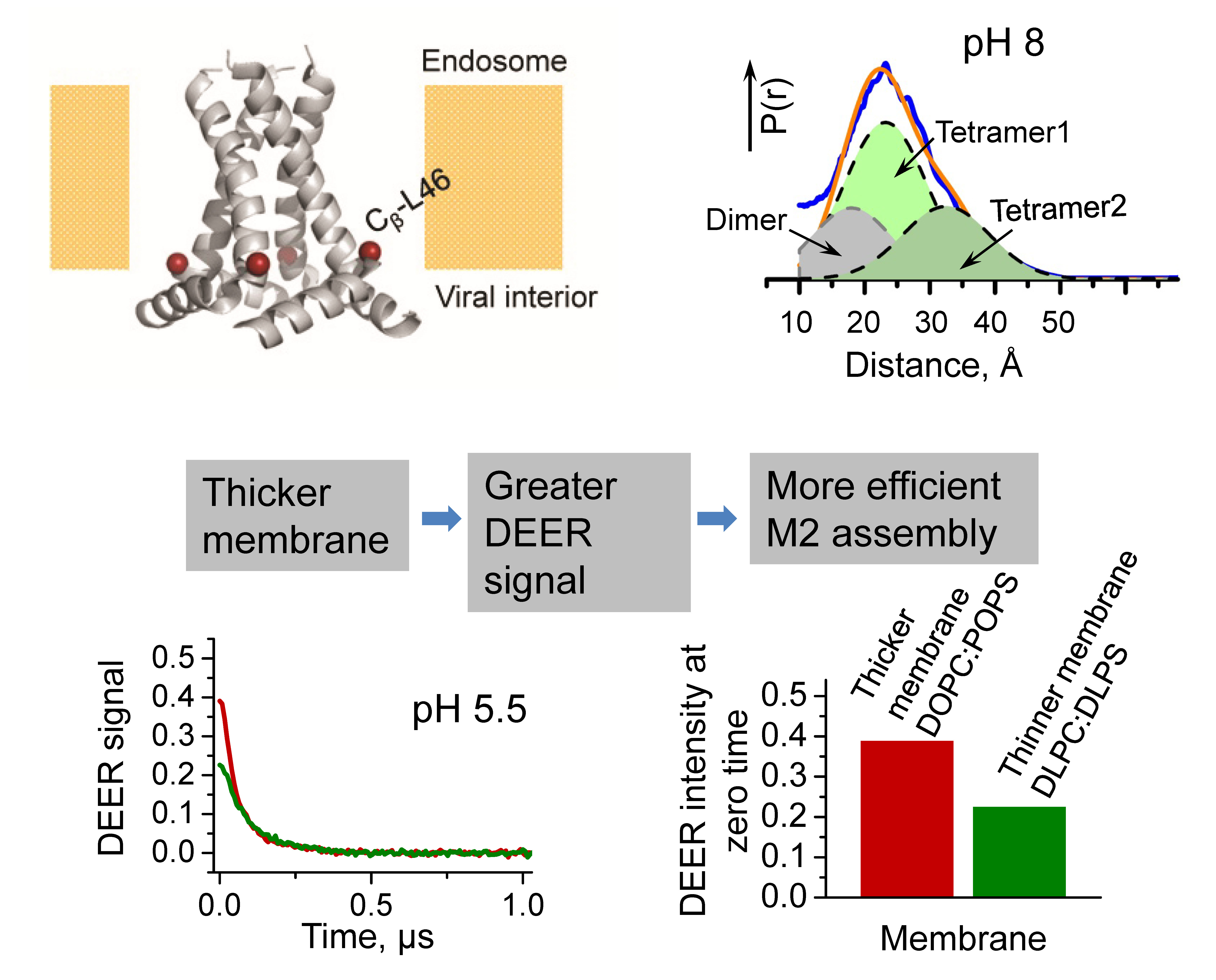
The influenza A M2 protein is single-pass transmembrane protein that assembles in a tetramer, forming a pH-activated proton channel (cf. Fig., upper left). It is essential for viral function. The tetramer of M2 transmembrane domain (M2TMD), residues 22-46, is the minimal unit for proton conductance. Currently, M2 inhibitors are the only existing anti-influenza A drugs. However, viral proliferation has been restored by efficient natural mutagenesis that prevents drug binding and inhibition. Therefore, new pharmaceuticals need to be developed in order to regulate M2 function and possibly suppress influenza viral infection. The high resolution structures, provided mostly by NMR and x-ray crystallography, have revealed several distinct conformations of M2TMD, suggesting a high structural variability. However, a more detailed knowledge about M2-membrane interactions needs to be acquired to aid drug development. Furthermore the mechanism of M2TMD assembly and structural alterations upon channel activation in lipid membranes is still not fully understood. We utilized pulsed ESR spectroscopy (DEER) with protein spin-labeling to study the M2TMD peptide (residues 21-49) in lipid membranes. A unique cysteine residue (L46C), introduced in M2TMD, was spin-labeled with MTSL. The peptide was reconstituted in membranes of different composition, i. e. zwitterionic or mixtures of zwitterionic and anionic lipids, as well as variable thickness. We measured inter-spin distances and their distributions (P(r), cf. Fig., upper right) between membrane-embedded M2TMD spin-labeled monomers in order to elucidate their assembly. Based on the modulation depth of DEER signals, that is the signal amplitude at zero evolution time (cf. Fig. lower left and right), and the reconstructed distance distributions, our results suggest that membrane-associated M2TMD is polydisperse and most likely exists as monomers, dimers and tetramers. We also found that the assembly and the structure of M2TMD oligomers are directly coupled to the membrane hydrocarbon thickness. Moreover, anionic lipids at physiological concentration are required for M2TM channel opening at low pH. Thus, our results contribute to the understanding of M2 properties and also to the fundamental knowledge about folding and assembly of transmembrane helices. Publication: E.R. Georgieva, P.P. Borbat, H.D. Norman, and J.H. Freed, Sci. Rep., 5, 11757 (2015); PMC4507135. |
|
|
|
|
Elka R. Georgieva, Peter P. Borbat, and Jack H. Freed (ACERT) Haley D. Norman (College of Engineering, Cornell University) |
|
|
|
About ACERT Contact Us |
Research |
Outreach |
ACERT is supported by grant 1R24GM146107 from the National Institute of General Medical Sciences (NIGMS), part of the National Institutes of Health. |
|||||
| ||||||||